Der Lactose-Abbau durch das lac-Operon
Auf der Seite über das lac-Operon haben wir schon Einiges über den Abbau von Lactose gelesen, vor allem, wie dieser Abbau reguliert wird. Betrachten wir diesen Lactose-Abbau nun einmal aus biochemischer Sicht:
Lactose ist ein Disaccharid und besteht aus je einem Glucose- und einem Galactose-Molekül.
Bei dem Abbau von Lactose wird das Molekül von E. coli zunächst in die beiden Monosaccharide Glucose und Galactose gespalten, bereits das ist schon ein energieaufwändiger Prozess.
Mit der Galactose kann die Bakterienzelle nicht viel anfangen, daher wird dieses Monosaccharid in Glucose umgewandelt - ebenfalls ein energieaufwändiger Prozess.
Am Ende haben wir also zwei Glucose-Moleküle vorliegen, die aus der Glucose gewonnen wurden - eines direkt, das andere durch Umwandlung der Galactose.
Die Glucose fließt nun direkt in die Glycolyse ein, dann in den Citratzyklus und schließlich in die Atmungskette.
Was geschieht nun, wenn man Bakterien gleichzeitig Lactose und Glucose als Nährstoff anbietet? Wie durch die obigen Ausführungen hoffentlich klar geworden ist, ist Glucose für die Bakterien wesentlich attraktiver als Lactose, da hier nichts mehr zerlegt werden muss. Es würde also durchaus Sinn machen, wenn die Bakterien zunächst die vorhandene Glucose abbauen und die auch anwesende Lactose ignorieren würden. Und genau das passiert auch:
In Anwesenheit von Glucose findet keine lac-Transkription statt, auch dann nicht, wenn genügend Lactose zur Verfügung steht.
Jetzt fragt sich natürlich jeder und jede: "Wie soll das denn funktionieren?"
Wenn Lactose anwesend ist, setzt sich doch die Lactose in das allosterische Zentrum des lac-Repressors, der ändert seine Konformation und löst sich vom Operator. Der Weg für die RNA-Polymerase wird frei, und die Transkription der drei Lactose abbauenden Enzyme beginnt.
Wenn Glucose-Moleküle in der Zelle vorhanden sind, muss irgendetwas passieren, was die Transkription der lac-Strukturgene verhindert. Es muss also ein zweites Kontrollsystem vorliegen. Ein zweiter Repressor vielleicht, der durch Glucose aktiviert wird? Oder ein anderer Mechanismus?
Mechanismus der Steuerung durch Glucose
In der Tat gibt es neben dem Kontrollsystem durch den lac-Repressor (negative Kontrolle, weil ja ein Repressor-Protein dafür verantwortlich ist) ein zweites Kontrollsystem, das den Lactose-Abbau reguliert. Diesmal handelt es sich aber um eine positive Kontrolle durch ein Aktivator-Protein.
Negative Kontrolle
Die Transkriptionsrate eines Gens oder eines Operons wird durch ein Repressor-Protein kontrolliert. Die Transkription ist nur in Abwesenheit des Repressors möglich. Sowohl eine Substratinduktion wie auch eine Endproduktrepression ist über einen solchen Repressor möglich.
Positive Kontrolle
Die Transkriptionsrate eines Gens oder eines Operons wird durch ein Aktivator-Protein kontrolliert. Die Transkription ist nur in Anwesenheit des Aktivators möglich bzw. wird durch den Aktivator stark gefördert. Sowohl eine Substratinduktion wie auch eine Endproduktrepression ist über einen solchen Repressor möglich.
Das Aktivator-Protein CAP
Neben Repressor-Proteinen, die die RNA-Polymerase blockieren, gibt es auch Aktivator-Proteine, welche die Polymerase aktivieren. Im Falle des lac-Operons heißt dieser Aktivator CAP. CAP ist die Abkürzung für "catabolic activator proteine".
Aufgabe
Begründen Sie, ob CAP aktiv oder inaktiv sein muss, wenn genügend Glucose zur Verfügung steht.
Wenn genügend Glucose zur Verfügung steht, sollen die lac-Enzyme nicht produziert werden, selbst dann, wenn viel Lactose vorhanden ist.
Der lac-Repressor hat sich in diesem Fall vom lac-Operator gelöst, die RNA-Polymerase könnte also durchaus eine Transkription der lac-Gene einleiten. In Anwesenheit von Glucose ist es aber sinnvoll, die Transkription nicht zu aktivieren oder sogar zu bremsen. Der lac-Aktivator darf also in Anwesenheit von Glucose nicht aktiv sein.
Umgekehrt heißt das: In Abwesenheit von Glucose ist der lac-Aktivator aktiv, setzt sich also an die RNA-Polymerase und aktiviert diese. Ist nun auch der Operator frei, weil Lactose zur Verfügung steht, kann die Transkription stattfinden.
Betrachten wir nun folgende vier Fallbeispiele:
| Glucose vorhanden | Glucose nicht vorhanden | |
| Lactose vorhanden |
CAP inaktiv |
CAP aktiv |
| Lactose nicht vorhanden |
CAP inaktiv |
CAP aktiv |
Eine Transkription der lac-Gene findet also nur dann statt, wenn
a) Lactose vorhanden ist, weil dann der lac-Repressor nicht am Operator sitzt und die Polymerase blockiert
b) und gleichzeitig keine Glucose vorhanden ist, weil dann der lac-Aktivator die Polymerase aktiviert.
Wer steuert die Aktivität von CAP?
Im Neurophysiologie-Kurs haben wir ionotrope und metabotrope Synapsen kennengelernt. Hier noch mal ein kurzer Überblick:
Bei den ionotropen Synapsen setzt sich der Neurotransmitter direkt in einen Ionenkanal der postsynaptischen Membran und öffnet diesen Kanal, so dass Ionen in die postsynaptische Zelle eindringen können.
Bei den metabotropen Synapsen dagegen setzt sich der Neurotransmitter an ein Rezeptor-Protein, das auf der Zellinnenseite mit einem G-Protein verbunden ist. Das G-Protein wiederum aktiviert eine Adenylatcyclase, und dieses Enzym stellt aus dem allgegenwärtigen ATP den second messenger cAMP her.
Dieser second messenger hat vielfältige Funktionen in der Zelle, er kann sich zum Beispiel in Ionenkanäle setzen und diese öffnen. Er kann aber auch bestimmte Enzyme aktivieren oder deaktivieren, die ihrerseits andere Enzyme (oder Transkriptionsfaktoren!) aktivieren bzw. deaktivieren.
Der CAP/cAMP-Komplex
Sie haben es schon geahnt, warum hätte ich sonst hier ausführlich über die Rolle des second messengers cAMP schreiben sollen? CAP hat ein allosterisches Zentrum, und in dieses Zentrum passt ein cAMP-Molekül hinein.
CAP ist nur dann aktiv, wenn dieses allosterische Zentrum durch cAMP besetzt ist. Nur dieser CAP/cAMP-Komplex kann als Aktivator arbeiten und die RNA-Polymerase zur Transkription der lac-Gene anregen.
Damit sind wir der Auflösung der Frage, wie Glucose die Transkription der lac-Gene verhindert, einen Schritt näher gerückt.
Offensichtlich muss die Anwesenheit von Glucose verhindern, dass sich cAMP in den CAP-Aktivator setzt und diesen aktiviert.
In Abwesenheit von Glucose kann sich das cAMP dagegen ungehindert in den CAP-Aktivator setzen und dieses aktivieren, so dass dieser die RNA-Polymerase aktivieren kann, um die lac-Gene zu transkribieren (falls nicht der lac-Repressor im Wege ist, weil die Lactose-Konzentration zu niedrig ist).
In der Tat kann die Anwesenheit von Glucose sich auf die cAMP-Konzentration auswirken. Die Bildung von cAMP wird nämlich unterdrückt, wenn viel Glucose im Außenmedium vorhanden ist. Bei geringen Glucose-Konzentrationen dagegen wird viel cAMP gebildet.
Fassen wir unsere Erkenntnisse wieder einmal zusammen:
Wenig Glucose → viel cAMP → CAP ist aktiv → Polymerase wird aktiviert → Transkriptionsrate ist hoch.
Viel Glucose → wenig cAMP → CAP ist passiv → Polymerase wird nicht aktiviert → Transkriptionsrate ist niedrig.
Glucose und cAMP
Der second messenger cAMP wird von dem Enzym Adenylatcyclase aus ATP hergestellt. ATP ist ja als universeller Energieträger der Zelle bekannt. Die Adenylatcyclase entfernt zwei Phosphatgruppen aus dem ATP-Molekül und schließt den Rest zu einem Ring. Daher auch der Name: zyklisches Adenosinmonophosphat oder cAMP.
Wie kann nun die Glucose-Konzentration die Aktivität der Adenylatcyclase beeinflussen? Eine entscheidende Rolle spielt das Glucose-Transportprotein der Bakterienzelle.
Im Fall des lac-Operons werden die Adenylatcyclasen durch die Glucose-Transportproteine gehemmt: Wenn im Zellaußenmedium eine hohe Glucose-Konzentration herrscht, so transportiert dieses Protein den Zucker in das Zellinnere und hemmt dabei die Adenylatcyclasen auf der Innenseite der Membran. So sinkt der cAMP-Spiegel im Zellinnern, und die Zahl der CAP/cAMP-Komplexe geht zurück. Die RNA-Polymerase wird nicht aktiviert kann die lac-Gene daher nicht oder nur mit geringer Effizienz transkribieren.
In Abwesenheit von Glucose ist dagegen der cAMP-Spiegel in der Zelle recht hoch, darum gibt es viele CAP/cAMP-Komplexe, die ihrerseits die RNA-Polymerase aktivieren. Die lac-Gene werden mit hoher Effizienz transkribiert, so dass Lactose abgebaut werden kann.
Fassen wir zusammen:
Wenig Glucose → kein Glucose-Transport → Adenylatcyclase wird nicht gehemmt → viel cAMP-Synthese → CAP/cAMP ist aktiv → RNA-Polymerase wird aktiviert → Transkriptionsrate ist hoch.
Viel Glucose → intensiver Glucose-Transport → Adenylatcyclase wird gehemmt → wenig cAMP-Synthese → CAP ist passiv → RNA-Polymerase wird nicht aktiviert → Transkriptionsrate ist niedrig.
Und das Ganze noch mal als Bild zusammengefasst:
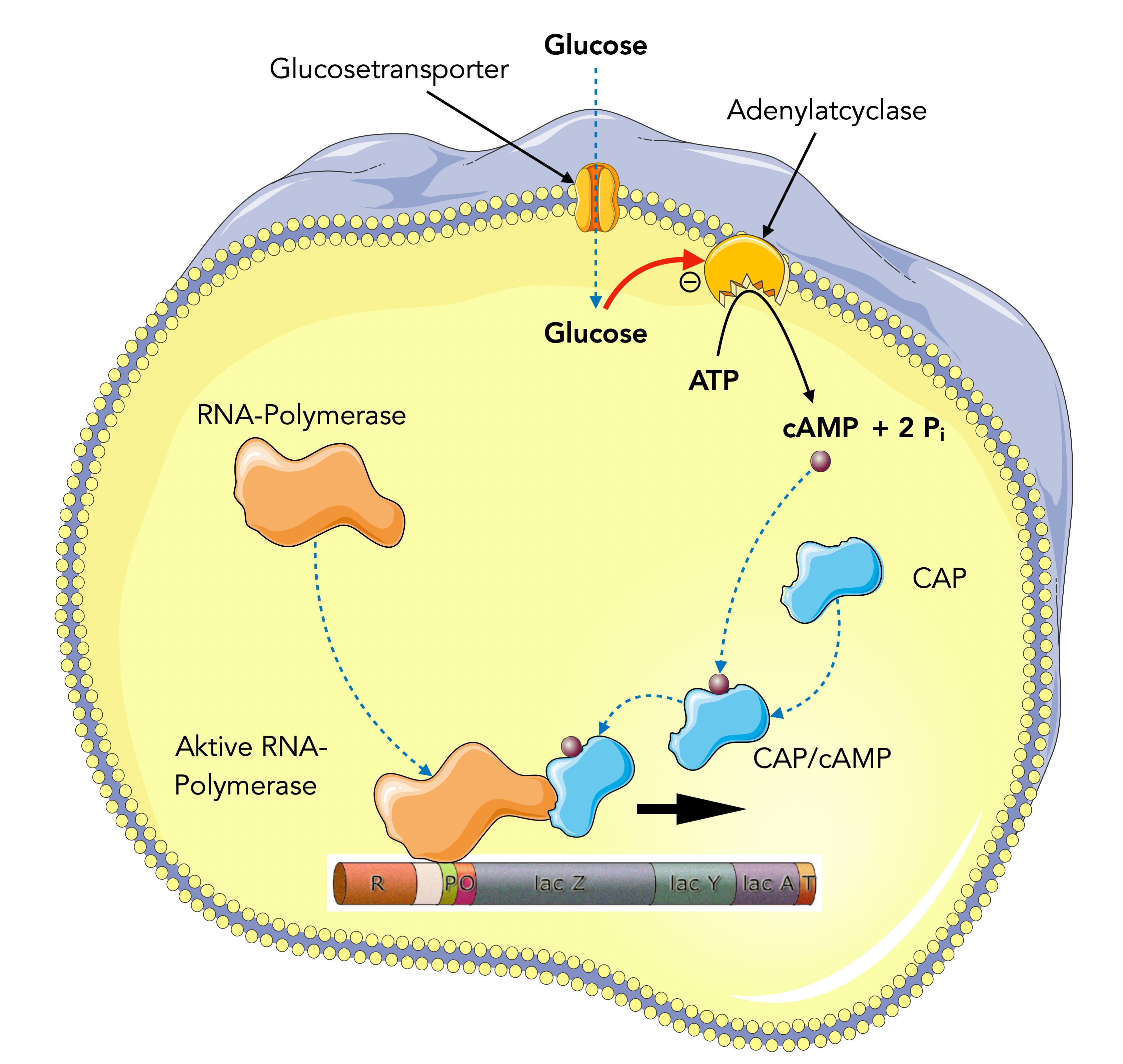
Einfluss von Glucose auf die Transkription des lac-Operons
Autor: Ulrich Helmich 2022. Das Bild wurde erstellt unter Verwendung von Symbolen von Servier Medical Art.
Servier Medical Art by Servier ist lizensiert unter einer Creative Commons Attribution 3.0 Unported License
Glucose hemmt die Lactose-Permease
Es gibt noch einen hemmenden Effekt von Glucose auf das lac-Operon. Der Transport von Glucose in das Zellinnere bewirkt eine Hemmung des Enzyms Lactose-Permease. Dieses Enzym wird ja von dem Gen lac Y des lac-Operons codiert. Wenn die Lactose-Permease gehemmt ist, kann keine Lactose mehr in das Zellinnere transportiert werden, und der lac-Repressor kann nicht deaktiviert werden. Die Transkription des lac-Operons bleibt also gehemmt, wenn Glucose anwesend ist.
CAP/cAMP fördert den Glucose-Transport
Und noch einen interessanten regulatorischen Effekt gibt es, der das lac-Operon betrifft. Der CAP/cAMP-Komplex aktiviert nicht nur das lac-Operon, sondern auch viele andere Gene. Unter anderem das Gen ptsG, das einen wichtigen Glucose-Transporter codiert.