Lernziele
Wenn Sie diese Seite durchgearbeitet haben, sollten Sie wissen
- was man unter der allopatrischen Artbildung versteht,
- in welchen Einzelschritten die allopatrische Artbildung abläuft,
- welche Rolle der Gründereffekt bei der allopatrischen Artbildung spielt,
- was der Unterschied zwischen Separation und Isolation ist,
- welche Bedeutung Isolationsmechanismen bei der allopatrischen Artbildung haben.
Schritt 1: Geographische Separation
Bei der allopatrischen Artbildung ist der erste und gleichzeitig wichtigste Schritt die geographische Separation von zwei Populationen einer Art.
Manchmal werden Populationen durch Naturereignisse in zwei Gruppen gespalten, in zwei Teilpopulationen.
Ein Fluss kann zum Beispiel plötzlich seinen Lauf ändern und dadurch das Siedlungsgebiet einer Kaninchengruppe zerteilen. Ein Gebirge kann sich auftürmen, ein Grabenbruch zerteilt eine Landschaft, oder aus einem großen See werden durch lange Trockenzeiten zwei oder drei kleinere Seen.
Aber auch auf andere, weniger spektakuläre Weisen können Populationen in zwei Teilpopulationen getrennt werden. Beispielsweise kann durch einen starken Sturm ein Teil eines Vogelschwarms abgetrennt und auf eine ferne Insel verweht werden. Dann tritt der sogenannte Gründereffekt auf.
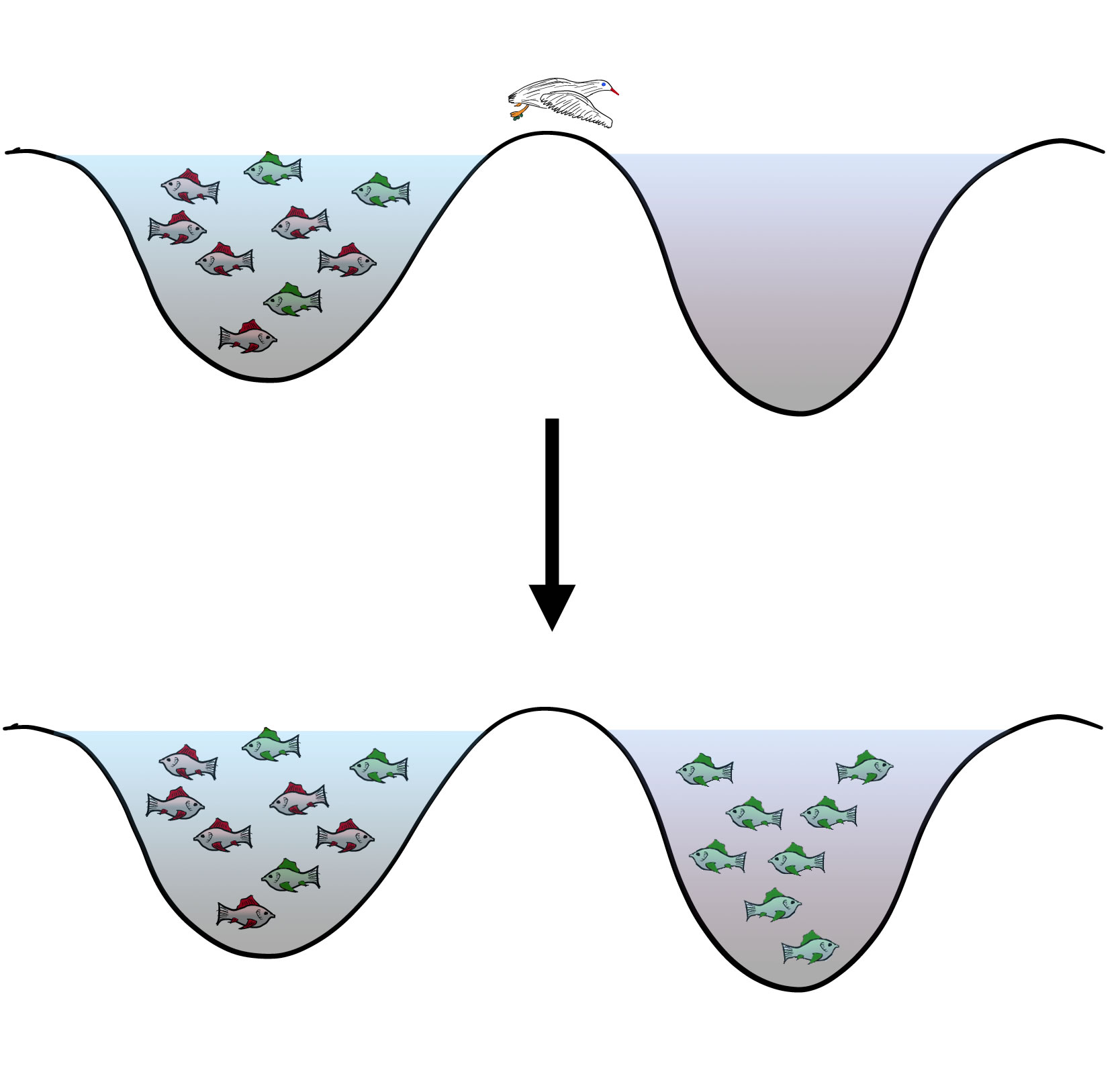
Ein Vogel trägt unfreiwillig Eier nur der grünen Fische in einen neuen See
Oder ein Wasservogel, zum Beispiel ein Schwan oder eine Ente, hält sich in einem See auf, in dem rote und grüne Exemplare einer bestimmten Fischart vorkommen. An seinen Füßen bleiben nun ein paar Eier der grünen Fische kleben. Der Vogel fliegt nun zum nächsten See, in dem diese Fischart noch nicht vorkommt. Dort entwickeln sich dann die Eier, und es entsteht eine neue Population dieser Art, die aber nur aus grünen Fischen besteht.
Entscheidend bei diesen Vorgängen ist: Eine Ursprungspopulation wird physikalisch in zwei Teilpopulationen getrennt, die sich von nun an unabhängig voneinander weiterentwickeln. Voraussetzung für diese unabhängige Weiterentwicklung der beiden Teilpopulationen ist allerdings eine ständige Unterbrechung des Genflusses zwischen den beiden Teilpopulationen, womit wir schon beim nächsten Schritt der allopatrischen Artbildung wären.
Schritt 2: Unterbrechung des Genflusses
Durch diese geographische bzw. physikalische Separation wird der Genfluss zwischen den neu entstandenen Teilpopulationen effektiv unterbrochen. Das heißt, die Individuen aus der einen Teilpopulation können sich nicht mehr mit den Individuen aus der anderen Teilpopulation fortpflanzen. Damit wird nach dem Biologischen Artkonzept bereits der Weg zu einer Artaufspaltung eingeleitet.
Wenn beispielsweise eine Ameisenart, die auf dem Boden nach Nahrung sucht, eine Nahrungsknappheit erlebt, kann es sein, dass sich ein Teil der Population entschließt, auf Bäumen nach Nahrung zu suchen. Sie erschließen sich so eine neue ökologische Nische. Die beiden Teilpopulationen sind jetzt zwar räumlich getrennt, aber nicht physikalisch oder geographisch. Jederzeit können die baumlebenden Ameisen wieder auf den Boden krabbeln und umgekehrt. Eine physikalische Separation liegt hier nicht vor, somit kann auch keine allopatrische Artbildung stattfinden.
Schritt 3: Unabhängige Entwicklung der beiden Gruppen
Wenn nun in der einen Teilpopulation eine Mutation auftritt, so kann diese sich nur innerhalb dieser Gruppe ausbreiten, falls sie neutral oder sogar vorteilhaft ist (schädliche Mutationen werden sich wohl kaum ausbreiten). Das Gleiche gilt für vorteilhafte oder neutrale Mutationen in der zweiten Teilpopulation. Selbst dann, wenn die Umweltbedingungen absolut gleich sind, entwickeln sich beide Teilpopulationen daher langsam aber sicher auseinander.
Würde noch ein Genfluss zwischen den beiden Teilpopulationen bestehen, so könnten sich beide Mutationen in beiden Gruppen ausbreiten, der Genpool beider Gruppen wäre einheitlich. Durch die räumliche Trennung aber wird dieser Genfluss unterbrochen, und die Mutationen können sich nur in der jeweiligen Gruppe ausbreiten. Da in jeder Gruppe mit der Zeit viele neue Mutationen entstehen, werden die beiden Teilpopulationen mit der Zeit immer unähnlicher.
Ursache für diese divergierende Evolution ist das zufällige Auftreten neutraler und vorteilhafter Mutationen in den beiden Teilpopulationen. Wesentlich schneller verläuft diese Entwicklung, wenn die beiden Gruppen unterschiedlichen Umweltbedingungen und damit unterschiedlichen Selektionsdrücken ausgesetzt sind.
Stellen wir uns einmal vor, bei den oben erwähnten Kaninchen würde die eine Teilpopulation in ein nördlich gelegenes kaltes Gebiet abgedrängt, in dem überdurchschnittlich viel Schnee fällt, und die andere Teilpopulation in ein südlich gelegenes Gebiet, in dem wenig Schnee fällt. In beiden Gruppen treten durch zufällige Mutationen immer wieder mal weiße Kaninchen auf.
Eine solche Mutation ist normalerweise nachteilig, weil weiße Kaninchen besser von Fressfeinden gesehen werden können. Es herrscht also ein Selektionsdruck in Richtung dunkler Fellfarbe. Daher sollte die Anzahl weißer Kaninchen in einer Population sehr sehr klein sein.
Allerdings haben weiße Kaninchen auf einem Boden, der mit Schnee bedeckt ist, einen Überlebensvorteil, denn sie werden von ihren Fressfeinden nicht mehr so gut erkannt wie normal gefärbte Artgenossen.
Wenn man dies berücksichtigt, könnte man also folgende Vorhersage für die beiden Kaninchenpopulationen treffen: In der südlichen Population wird es kaum weiße Kaninchen geben, weil die weiße Fellfarbe nachteilig ist. In der nördlichen Population dagegen werden die weißen Kaninchen immer häufiger, weil die weiße Fellfarbe hier einen Überlebensvorteil bedeutet.
Eine ähnliches Phänomen hat man ja bei den berühmten Birkenspannern festgestellt, Schmetterlingen, die auf der weißen Rinde von Birken sitzen. Die meisten Exemplare sind weiß gefärbt, es gibt aber auch dunkle Individuen. In stark industrialisierten Gebieten Englands sind die Birken dunkler als üblich, und tatsächlich fand man in solchen Gegenden auch überdurchschnittlich viele Exemplare der dunklen Birkenspanner. Vermutlich werden die dunklen Birkenspanner auf den dunklen Birkenrinden weniger gut von ihren Fressfeinden gesehen.
Schritt 4: Rassenbildung
Die Unterschiede zwischen den Teilpopulationen werden im Laufe der Zeit immer stärker, bis sie sogar biologischen Laien auffallen. Jetzt könnte man sagen, dass aus den Teilpopulationen unterschiedliche Rassen geworden sind.
Rassen sind Varianten ein- und derselben Art, die sich in bestimmten Merkmalen voneinander unterscheiden. Man denke nur an die verschiedenen Hunde- oder Katzenrassen. Da Individuen unterschiedlicher Rassen immer noch der gleichen Art angehören, können sie sich auch miteinander fruchtbar fortpflanzen.
Entfällt also die geographische Barriere, die bisher die beiden Teilpopulationen bzw. Rassen getrennt hat und kommen die beiden Rassen wieder zusammen, dann steht einer Hybridisierung (genetische Vermischung) nichts im Wege. Durch den kontinuierlichen Genfluss, der dann einsetzt, verschwinden die unterschiedlichen Merkmale wieder, die die beiden Rassen unterscheiden.
Ein klassisches Beispiel für eine allopatrische Artbildung, die mitten in Schritt 4 stehen geblieben ist, sind die beiden europäischen Krähenarten Rabenkrähe und Nebelkrähe. Während der letzten Eiszeit kam es durch Gletscherbildung in Europa zu einer Trennung der Krähenpopulation. Die beiden Teilpopulationen entwickelten sich unterschiedlich, es entstanden die Rabenkrähen und die Nebelkrähen. Nach dem Ende der Eiszeit, als die Gletscher wieder zurückwichen, kamen sich die beiden Rassen wieder näher - und die Fortpflanzung war noch möglich. Die Mischformen zwischen Raben- und Nebelkrähe sind fruchtbar.
Schritt 5: Isolationsmechanismen
Nach vielen weiteren Generationen der getrennten Entwicklung sind die Unterschiede zwischen den beiden Rassen so groß geworden, dass eine fruchtbare Fortpflanzung zwischen Vertretern und Vertreterinnen der beiden Rassen nicht mehr möglich ist. Die Gründe dafür, dass keine fruchtbaren Nachkommen mehr gebildet werden können, sind vielfältig. Aber auf jeden Fall wird verhindert, dass sich die beiden Rassen nicht mehr fruchtbar fortpflanzen können, sie sind quasi voneinander fortpflanzungstechnisch isoliert, und daher fasst man alle Faktoren, die zu dieser Isolation führen, als Isolationsmechanismen zusammen. Einzelheiten dazu finden sich auf der Spezialseite "Isolationsmechanismen".
Ein Beispiel aus der heimischen Vogelwelt soll das illustrieren. Grünspechte und Grauspechte sind vor ca. 100.000 Jahren aus einer gemeinsamen Mutterart entstanden. Gletscher der Eiszeit trennten die Population Europas in zwei Teilgruppen, eine westliche und eine östliche. Durch allopatrische Artbildung entstanden zunächst zwei unterschiedliche Rassen. Als die Spechte nach dem Zurückweichen der Gletscher in Mitteleuropa wieder zusammen kamen, konnten sie sich nicht mehr untereinander fortpflanzen.
Grünspechte fressen hauptsächlich Ameisen und halten sich daher fast ausschließlich auf dem Boden auf. Grauspechte dagegen picken in der Rinde von morschen Bäumen nach Insekten.
Die unterschiedliche ökologische Nischenbildung bei den Grün- und Grauspechten könnte einer der Gründe dafür sein, dass sie sich nicht mehr untereinander fortpflanzen.
Ob sich die Tiere jetzt noch fortpflanzen können, wenn man sie unter künstlichen Bedingungen gemeinsam hält, ist eine andere Frage. Sollte das der Fall sein, müsste man sagen, dass beide Tiergruppen noch der selben Art angehören. Wenn es in der Natur aber de facto zu keiner Fortpflanzung kommt, weil unterschiedliche ökologische Nischen besetzt sind, könnte man eigentlich schon von zwei verschiedenen Arten sprechen. Die Grenzen zwischen Rassen und Arten sind sehr fließend, wie dieses Beispiel zeigt.
Schritt 6: Artbildung
Wenn sich Individuen zweier Teilpopulationen oder Rassen nicht mehr fruchtbar miteinander fortpflanzen können, hat man es nach dem biologischen Artbegriff mit zwei verschiedenen Arten zu tun. Vertreter verschiedener Arten können keine fruchtbaren Nachkommen mehr hervorbringen. Ein Beispiel hierfür sind Pferd und Esel. Zwar kann sich ein Pferd mit einem Esel paaren, doch sind die Nachkommen - Maulesel und Maultier - steril.
Die beiden Arten, die aus der ursprünglichen Population bzw. aus der Mutterart hervorgegangen sind, bezeichnet man auch als Schwesterarten oder auch Geschwisterarten. Grünspecht und Grauspecht sind ein bekanntes Beispiel für solche Geschwisterarten. Da kein Genfluss zwischen den Geschwisterarten mehr besteht, auch wenn sie inzwischen wieder im gleichen Lebensraum vorkommen, kann die Evolution der beiden Arten weiterhin getrennte Wege gehen, und die Unterschiede zwischen den beiden Arten werden im Laufe der Zeit immer größer.
Abituraufgabe "Flussdelfine"
In der NRW-Abituraufgabe "Die Evolution der Amazonas-Flussdelfine" für den Grundkurs 2015 findet sich ein sehr schönes Beispiel für allopatrische Artbildung durch geographische Separation. Die drei Unterarten des Flussdelfins kommen in drei verschiedenen Gebieten Südamerikas vor, die durch zwei geographische Barrieren voneinander getrennt sind; die eine Barriere ist ein unüberwindliches Hindernis (Stromschnellen), die andere Barriere ist nicht ganz so gravierend; es handelt sich hier um eine "Schwarzwasserregion" im Flusssystem, die durch Humusmaterial dunkel gefärbt und leicht angesäuert ist. Diese Region wird von den Delfinen gemieden, aber vereinzelt findet man doch Exemplare beider Unterarten, so dass ein minimaler Genfluss noch besteht.